Ferroptosis—a previously unknown form of controlled cell death with significant similarities to oxytosis*—was identified just seven years ago. This lethal event does not carry the traditional hallmarks of necroptosis or apoptosis (e.g., organelle swelling, chromatin condensation, loss of membrane integrity). Instead, cells undergoing ferroptosis have only subtle differences in their morphological features, like smaller-than-normal cell volume and increased mitochondrial membrane density. The biochemistry of the process is different, too. Ferroptosis results from iron-dependent lipid peroxide accumulation. Iron greatly accelerates lipid peroxidation of fatty acids. When these lipid reactive oxygen species (ROS) exceed the capacity of a cell’s antioxidant system, the oxidative stress damages proteins, nucleic acids, and lipids. Thus, oxidized lipids serve as lethal signals to initiate a reaction leading to the removal/recycling of the damaged material.
Three key elements are needed to trigger the ferroptotic pathway:
1) Polyunsaturated long-chain fatty acids (PUFAs) stored in phospholipid membranes
2) Redox-active iron
3) A defective lipid peroxide repair system.
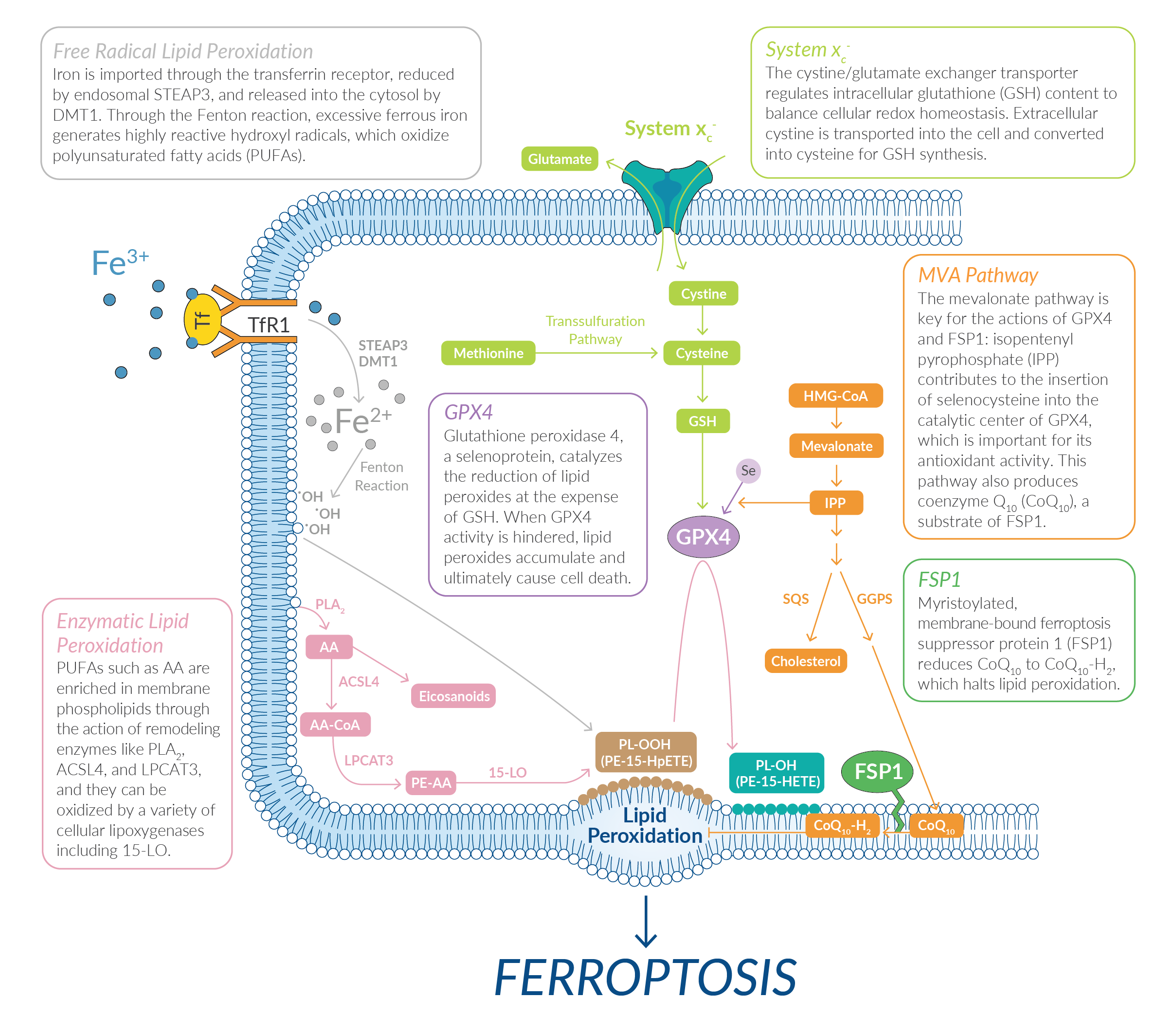
Several classes of small molecules can also trigger this process by targeting key proteins and/or related metabolic pathways (Figure 1).
Figure 1: The ferroptotic pathway can be triggered by several different classes of small molecules.
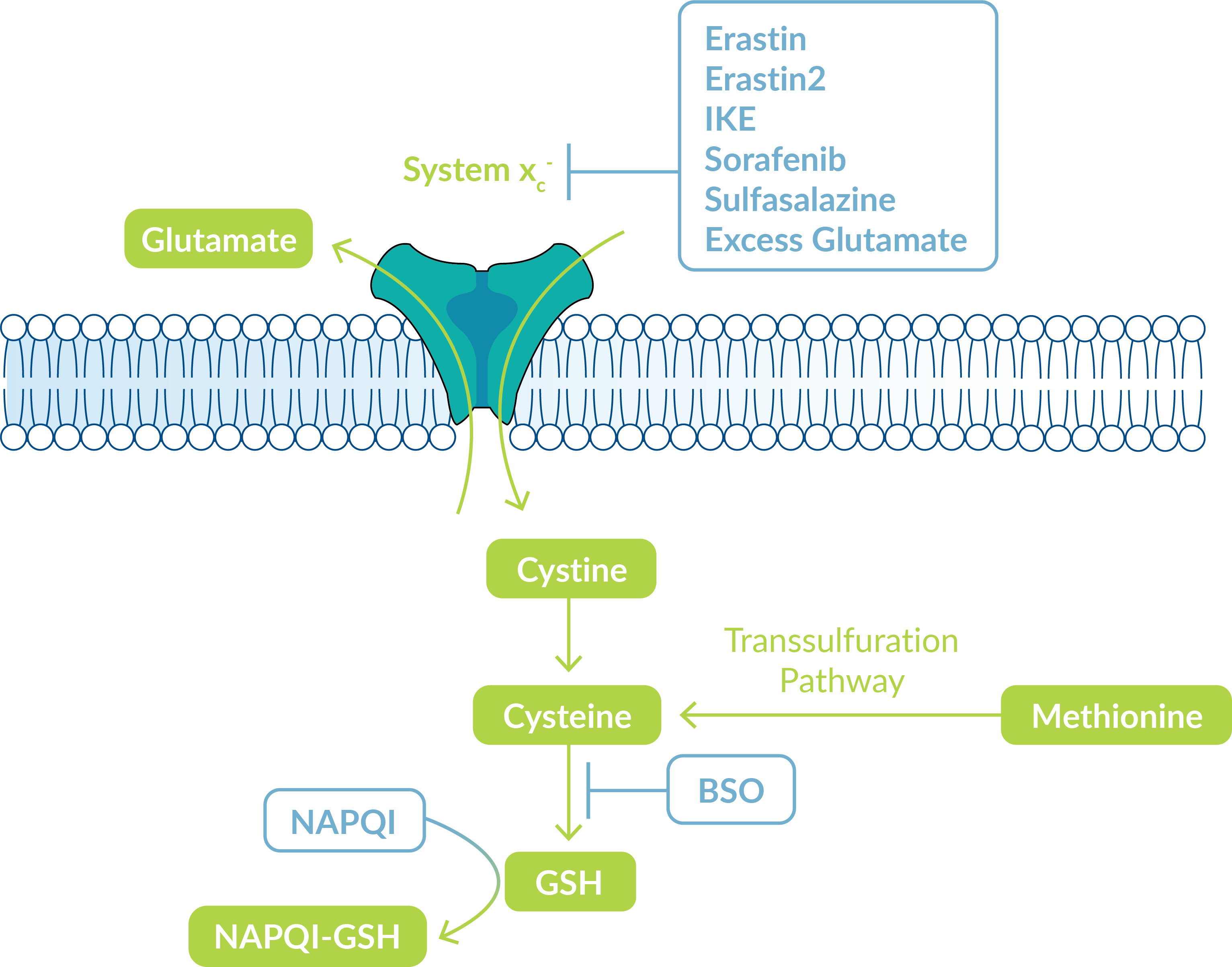
Certain ferroptosis inducers target the cystine/glutamate exchange transporter, system xc-, which regulates intracellular glutathione content to balance cellular redox. System xc- transports intracellular glutamate to the extracellular space while transporting extracellular cystine into the cell (Figure 2).
Figure 2: System XC- controls GSH synthesis.
All available cystine is then transformed into cysteine for glutathione (GSH) synthesis. Under conditions of cysteine insufficiency, methionine can be converted to cysteine through the transsulfuration pathway to ultimately supplement the cysteine pool. GSH is highly critical for protecting cells from damage caused by oxidative stress. Cells undergoing ferroptosis have very low levels of GSH and a disrupted oxidation-reduction balance. To study this process, Cayman Chemical offers several ferroptosis inducers that interfere with GSH production. Assay kits are also available to measure GSH as well as GSH reductase activity in cells.
| Ferroptosis Inducers of Cayman Chemical | Mode of action |
| Erastin | Inhibits uptake of cystine through system xc- |
| Erastin2 | Inhibits the system xc- cystine/glutamate transporter |
| Imidazole Ketone Erastin (IKE) | Metabolically stable inhibitor of system xc-; potentially suitable for in vivo applications |
| Sulfasalazine | A prodrug of 5-aminosalicylic acid that inhibits cystine uptake |
| Sorafenib | Inhibits cystine uptake, interfering with GSH production |
| L-Buthionine-(S,R)-Sulfoximine (BSO) | Depletes GSH by inhibiting γ-glutamylcysteine synthetase, the rate-limiting enzyme for GSH synthesis |
| N-Acetyl-4-benzoquinone imine (NAPQI) | Acetaminophen metabolite that conjugates with GSH during clearance; at toxic doses depletes available GSH liver reserves |
| Decreases intracellular glutathione levels |
*Products are available in different sizes.
| Glutathione Assays of Cayman Chemical | Glutathione Enzyme Assays / Antibodies of Cayman Chemical |
| Glutathione Assay Kit | Glutathione ethyl ester |
| Glutathione Cell-Based Detection Kit (Blue Fluorescence) | L-Glutathione, reduced |
| Glutathione Reductase Assay Kit | Glutathione S-Transferase Polyclonal Antibody |
| Glutathione S-Transferase Polyclonal FITC Antibody |
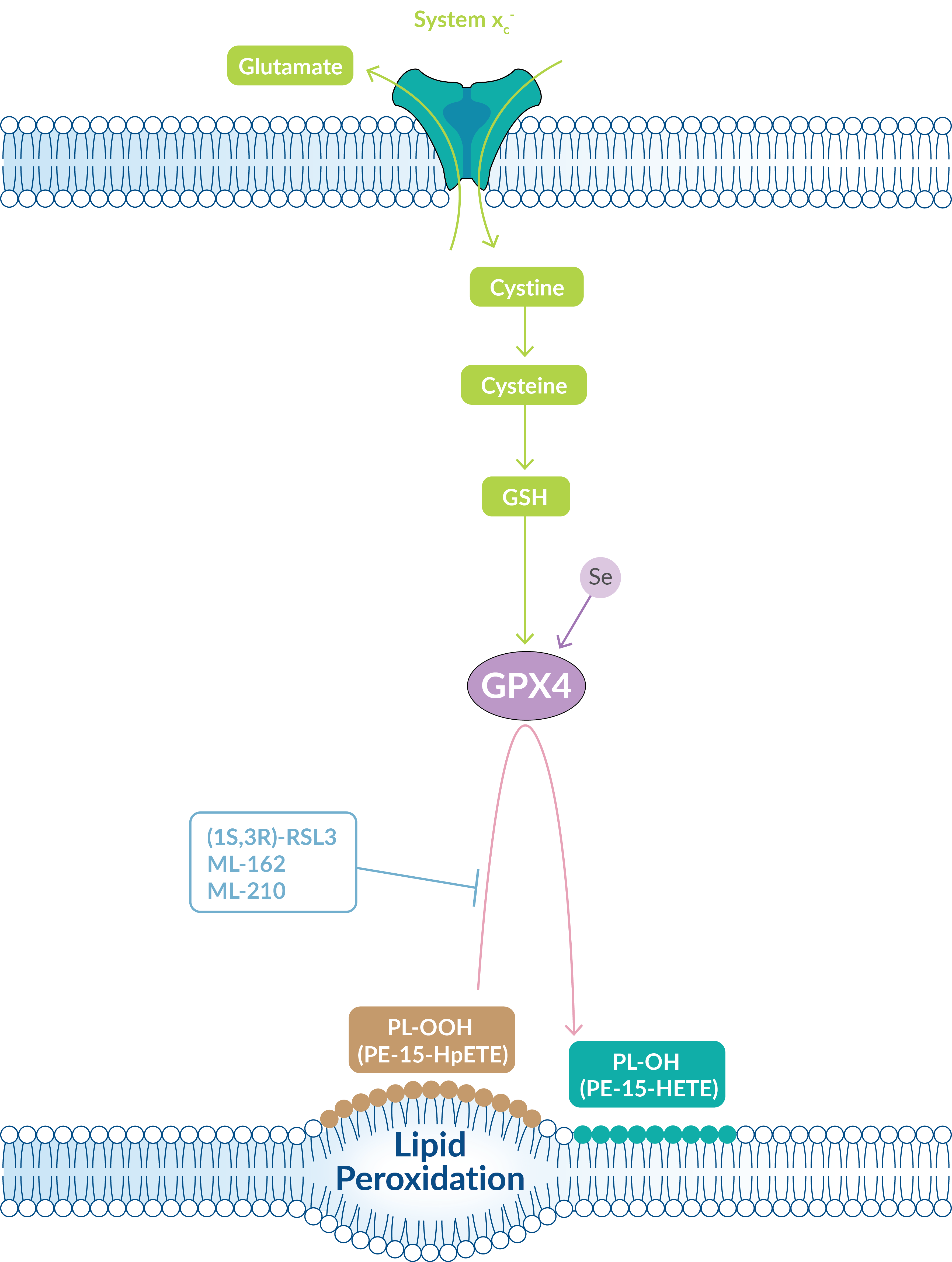
Other ferroptosis inducers directly target glutathione peroxidase 4 (GPX4), a lipid repair enzyme downstream of GSH that works to limit lipid peroxide accumulation (Figure 3). Thus, when the GSH pool is depleted, GPX4 activity is also reduced.
Figure 3: GPX4 functions downstream of GSH to repair lipid peroxidation.
GPX4 is a selenoprotein that functions inside biological membranes as a phospholipid hydroperoxidase. It catalyzes the reduction of lipid peroxides at the expense of reduced glutathione. When GPX4 activity is hindered, lipid peroxides accumulate and ultimately cause cell death. Cayman Chemical offers several direct suppressors of GPX4 activity that can be used to induce ferroptosis. These compounds work by covalent interaction with the selenocysteine active site of GPX4. A GPX4 antibody and assay kit are also available to study this enzymatic activity.
| Ferroptosis Inducers | Mode of Action |
| (1S,3R)-RSL3 | Directly binds the selenocysteine active site of GPX4 |
| ML-162 | Inhibits GPX4; more potent, selective than (1S,3R)-RSL3 |
| ML-210 | Inhibits GPX4 to induce ferroptosis |
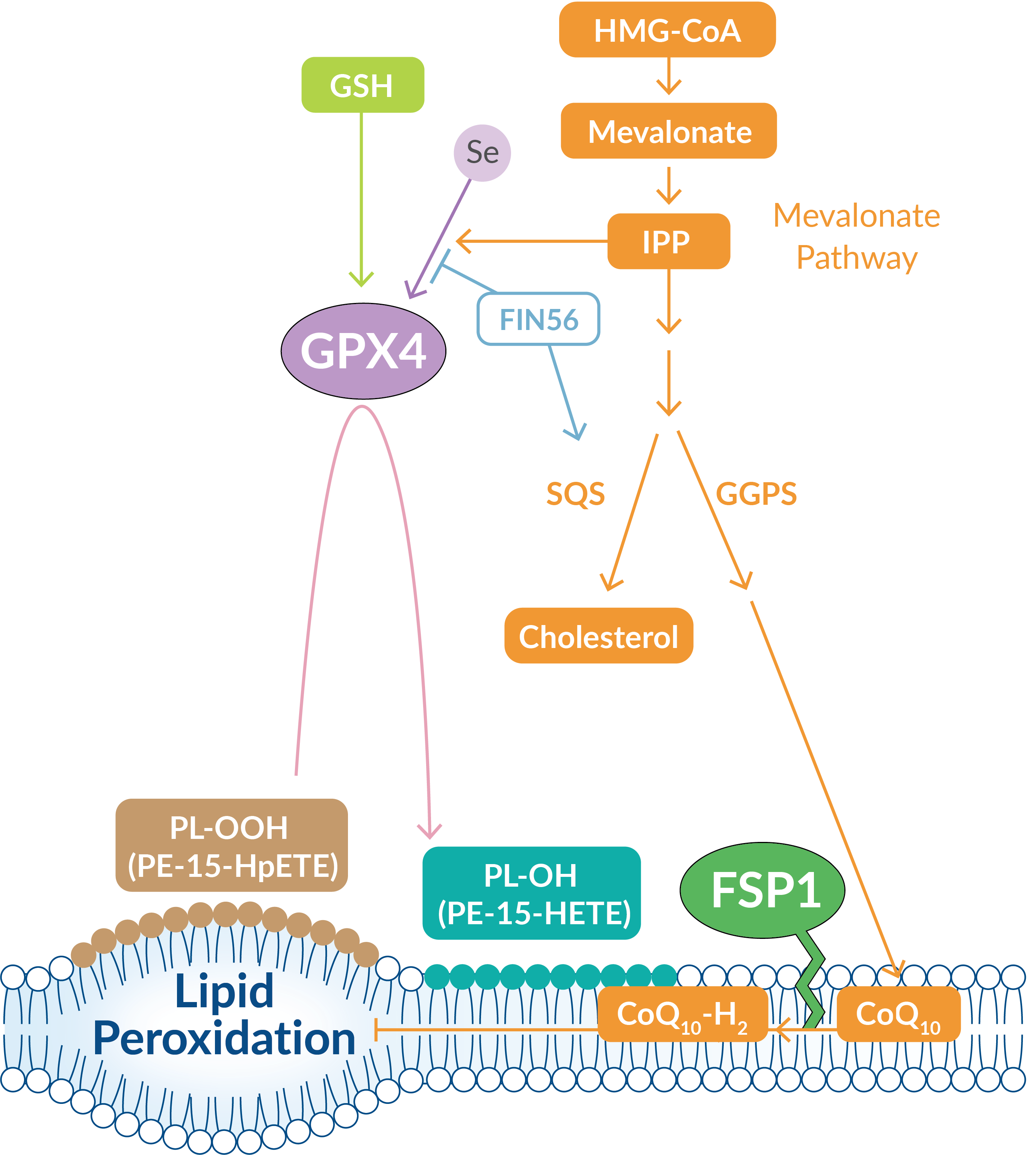
Dysregulation of lipid metabolism is also associated with ferroptosis. For instance, chemical activation of squalene synthase (SQS), an enzyme involved in cholesterol synthesis, suppresses the formation of several important non-sterol products in the mevalonate (MVA) pathway (Figure 4). One such product that is suppressed is coenzyme Q10 (CoQ10), an important free-radical scavenging antioxidant.
Ferroptosis is suppressed when a myristoylated version of ferroptosis suppressor protein 1 (FSP1) is recruited to the plasma membrane where it can reduce CoQ10 to a lipophilic antioxidant that traps lipid peroxides.
Figure 4. The MVA pathway plays important roles in the production of CoQ10 and the maturation of the GPX4 protein.
The MVA pathway also plays a key role in GPX4 maturation, as isopentenyl pyrophosphate (IPP) contributes to the complicated insertion of selenocysteine into the catalytic center of GPX4, which is important for its antioxidant activity. Cayman Chemical offers a ferroptosis inducer that triggers the production of ROS likely through a dysregulated MVA pathway.
| Ferroptosis Inducers of Cayman Chemical | Mode of Action |
| FIN56 | Reduces expression of GPX4 protein; also binds to and activates squalene synthase in a GPX4 degradation-independent manner |
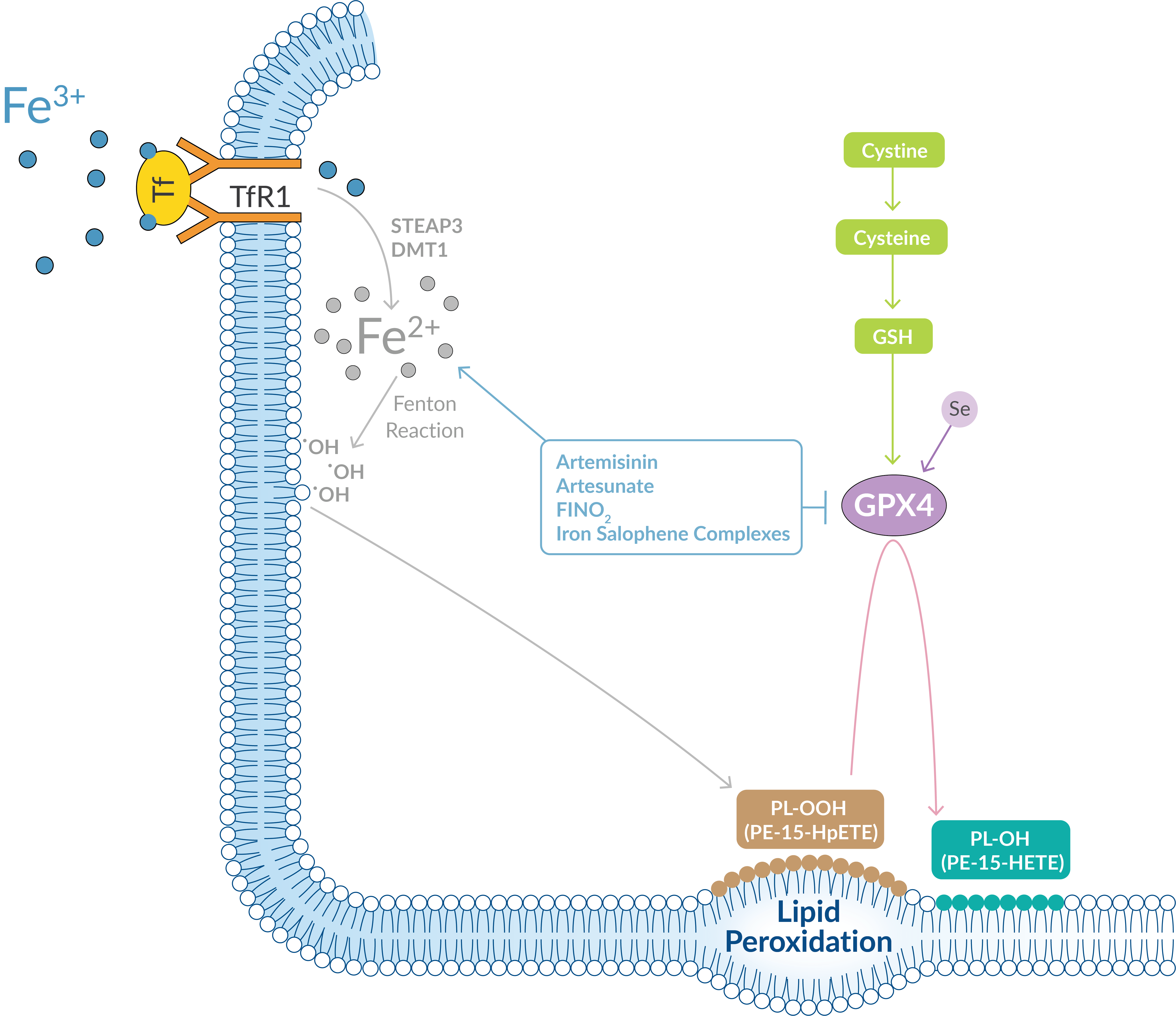
Excessive iron metabolism contributes to ferroptosis by producing oxidative stress. Circulating iron, in the form of ferric iron (Fe3+) bound to transferrin, is transported into cells through the membrane-bound transferrin receptor 1 (TfR1). Within endosome compartments, Fe3+ is reduced to ferrous iron (Fe2+) and released into a labile iron pool in the cytoplasm via the divalent metal transporter 1 (DMT1) (Figure 5). Extra iron is stowed in ferritin, an iron storage protein complex.
Figure 5. Excess iron leads to the production of ROS by the Fenton reaction.
Any increases in iron uptake or reduced capacity for iron storage contributes to iron overload and the potential to generate highly reactive hydroxyl radicals through the Fenton reaction. These radicals can oxidize PUFAs in lipid membranes, creating lipid hydroperoxides. Cayman Chemical offers several endoperoxides that act as ferroptosis inducers by oxidizing Fe2+, which promotes the production of lipid ROS.
| Ferroptosis Inducers | Mode of Action |
| Artemisinin | Iron(II) oxide-reactive endoperoxide that generates ROS |
| Artesunate | Iron(II) oxide-reactive endoperoxide that generates ROS |
| Chlorido[N,N'-disalicylidene-1,2-phenylenediamine]iron(III) | Metal complex that generates lipid-based ROS |
| FINO2 | Oxidizes iron, resulting in the loss of GPX4 enzymatic activity |
| Fluorescent Iron Indicator of Cayman Chemical | Description |
| Phen Green SK diacetate | A green fluorescent heavy metal indicator |
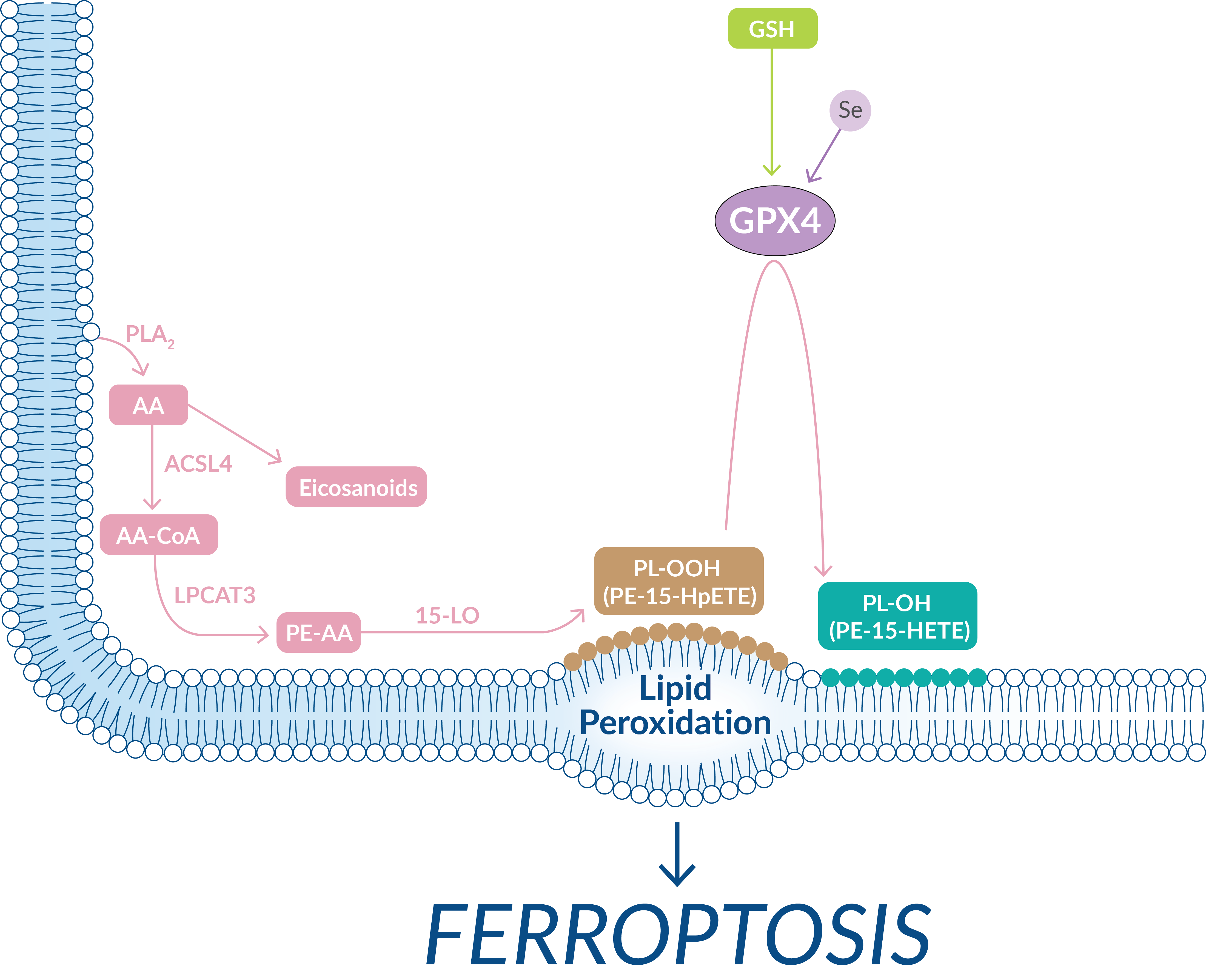
PUFAs are highly susceptible to oxidative damage through both non-enzymatic and enzymatic processes. PUFAs possess bis-allylic protons that are prone to hydrogen atom abstraction, which leads to the production of alkyl radicals that readily react with molecular oxygen to produce peroxyl radicals. These peroxyl radicals react with other PUFAs, creating a chain reaction of lipid peroxidation. In the course of ferroptosis, acyl-CoA synthetase long-chain family member 4 (ACSL4), lysophosphatidylcholine acyltransferase 3 (LPCAT3), and lipoxygenases (LOs) are the key drivers in the formation of oxidized PUFAs (Figure 6).
Figure 6. Membrane-bound lipid peroxidation, driven by LOs and unchecked by GPX4, triggers ferroptosis.
Arachidonic acid (AA) that is not converted to signaling oxylipins is conjugated to coenzyme A by ACSL4. LPCAT3 then catalyzes the esterification of AA into phosphatidylethanolamine (PE) within the phospholipid membrane. LOs, particularly 15-LO, which normally uses free PUFAs as substrates, is then capable of oxidizing PE-AA and generating hydroperoxy-PEs (e.g., PE-15-HpETE). If GPX4 inadequately reduces hydroperoxy-PEs, their accumulation will serve as a lethal signal to trigger ferroptosis. The release of 5-HETE, 11-HETE (but not 12-HETE), and 15-HETE has been observed following ferroptotic stimuli and treatment of cells with 5-HpETE, 12-HpHETE, and 15-HpHETE accelerates the ferroptotic process. Cayman Chemical scientists are experts in building the tools needed to study how oxidized lipids are involved in the ferroptotic process. This includes providing active LO enzymes as well as oxidized lipid standards that can be quantified as an index of lipid peroxidation.
| Lipoxygenases* of Cayman Chemical | Oxidized Lipid Mediators of Cayman Chemical |
| 5-Lipoxygenase (human recombinant) | 5(S)-HETE | 5(S)-HETE-d8 | 5(S)-HpETE |
| 12-Lipoxygenase (platelet-type, mouse recombinant) | 11(S)-HETE | 12(S)-HETE | 12(S)-HpETE |
| Lipoxygenase from Glycine max (soybean) - Purified | 15(S)-HETE | 15(S)-HETE-d8 | 15(S)-HpETE |
| 15-Lipoxygenase-2 (human recombinant) | (S)-HETE HPLC Mixture | Hydroperoxy HPLC Mixture |
| 15(S)-HETE ELISA Kit |
*Arachidonic Acid Alkyne can be used in click chemistry reactions to identify targets of LO-derived lipid ROS
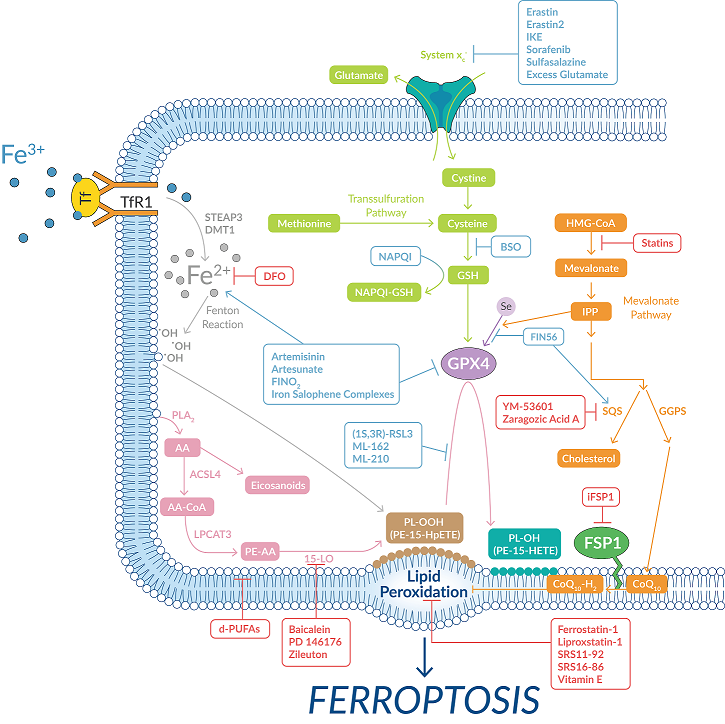
The presence of ferroptosis can be confirmed by looking at whether cell death is prevented by pertinent inhibitors and by measuring lipid peroxides. Cayman Chemical offers inhibitors for each of the major nodes within the ferroptotic pathway (Figure 7).
Figure 7. Targeting key points within the ferroptotic pathway through selective inhibition.
| Noted Ferroptosis Inhibitors of Cayman Chemical | Mode of Action |
| Ferrostatin-1 | Traps acyl chain-carrying peroxyl radicals in lipid bilayers |
| Ferrostatin-1 Diyne | A ferroptosis inhibitor with a diyne tag for localizing ferrostatin-1 in cells |
| UAMC-3203 | More stable, readily soluble ferrostatin-1 analog |
| iFSP1 | Inhibits FSP1, suppressing ferroptosis independent of glutathione activity |
| Liproxstatin-1 | Traps acyl chain-carrying peroxyl radicals in lipid bilayers |
| SRS11-92 | Inhibits ferroptotic cell death induced by erastin |
| SRS16-86 | Inhibits ferroptosis increasing GPX4, system xc- transport, and GSH |
| BHT, α-Tocotrienol, Trolox, Coenzyme Q10, Idebenone, Ebselen | Lipophilic antioxidants |
| Deferasirox, Deferiprone, Deferoxamine (mesylate)(DFO), Ciclopirox | Iron chelators |
| Cycloheximide | Protein synthesis inhibitor |
| YM-53601, Zaragozic Acid A | SQS inhibitors |
| TOFA | Acetyl-CoA carboxylase inhibitor |
| Statins | Inhibits HMG-CoA reductase, which catalyzes the rate limiting step of the MVA pathway |
| d-PUFAs | Deuterated PUFAs with deuterium ions in place of bis-allylic hydrogens slows radical generation |
| Baicalein, Nordihydroguaiaretic Acid, PD 146176, Zileuton | Lipoxygenase inhibitors |
| K252a,7-oxo Staurosporine | Phosphorylase kinase inhibitors |
| Thiazolidinediones | Inhibits the acyl-CoA synthetase essential for arachidonic acid metabolism |
Cayman Chemical also provides several convenient methods to detect the presence of lipid ROS. This includes assays for direct quantification of lipid hydroperoxides as well as assays that quantify end-product reactive aldehydes, such as malondialdehyde or 4-hydroxy nonenal (4-HNE).
| Lipid ROS Assays of Cayman Chemical | Mechanism |
| Lipid Hydroperoxide (LPO) Assay Kit | Measures hydroperoxides directly using the redox reactions with ferrous ions |
| Lipid Hydroperoxide (LPO) Assay Kit (96 well) | |
| TBARS (TCA Method) Assay Kit | Measures malondialdehyde, a byproduct of lipid peroxidation |
| TBARS Assay Kit | |
| DHN-MA EIA Kit | Measures a metabolite of 4-HNE, a byproduct of lipid peroxidation |
| ROS Detection Cell-Based Assay Kit (DHE) | Measure superoxide and hydrogen peroxide levels in living cells |
| Lipid ROS Probes of Cayman Chemical | Mechanism |
| 4-hydroxy Nonenal Alkyne | 4-HNE modified for click chemistry |
| DPPP | Fluorescent probe for detection of hydroperoxides |
| Lipid Peroxidation Products of Cayman Chemical | Deuterated Standards of Cayman Chemical |
| 4-hydroxy Nonenal | 4-hydroxy Nonenal-d3 |
| 4-hydroxy Nonenal Glutathione (trifluoroacetate salt) | 4-hydroxy Nonenal Glutathione-d3 (trifluoroacetate salt) |
| 4-hydroxy Nonenal Mercapturic Acid | 4-hydroxy Nonenal Mercapturic Acid-d3 |
| 4-hydroperoxy 2-Nonenal | |
| 4-oxo-2-Nonenal | 4-oxo-2-Nonenal-d3 |
While important for controlling damage brought on by the oxidative stress response, ferroptosis has also been implicated in several pathological conditions in the brain, kidney, liver, and heart. This indicates a necessity for the development of therapeutics to inhibit the process. Indeed, much more work must be done to fully understand the underworking of the ferroptotic pathway. With expertise in bioactive lipid synthesis and protein, antibody, and assay development, Cayman Chemical and Sanbio are here to help in this endeavor by providing much needed research tools.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}